
ADVERTISEMENTS:
The following points highlight the top twelve experiments on growth regulators in plants. Some of the experiments are: 1. Demonstration of the Effect of Auxin on Growth 2. Demonstration of Effect of Kinetin on Growth 3. Demonstration of effect of Gibberellin on Growth 4. Inhibition of Protoplasmic Movement 5. Inhibition of Water Uptake 6. Inhibition of Transpiration and Others.
Experiment # 1
Demonstration of the Effect of Auxin on Growth:
(a) On root growth and stem growth:
ADVERTISEMENTS:
Experiment:
0.25 % lanolin paste (wool fat) of auxin, viz., indole acetic acid (IAA) or indole butyric acid (IBA) or naphthalene acetic acid (NAA) is prepared by mixing 1 mg of auxin to 400 mg of lanolin.
A small quantity of this paste is applied to one side of some young stems of sunflower, tomato or tobacco or other suitable plants and to some other plants just at the base of the stem in the form of a girdle or ring. After a few days observations are made.
Observation:
ADVERTISEMENTS:
Bending of stem towards the untreated region of the stem is observed. In case of plants where an auxin girdle has been made root initials or even rootlets have been found to come out.
Inference:
Auxin increases markedly the plasticity of the walls of elongating cells and thus reviving the stage of elongation. Greater elongation of the cells occurs at the auxin treated region of the stem then the untreated region causing the bending of the stem.
It is the property of auxin to stimulate the initiation of cell division and rate of formation of roots when applied in lanolin paste to the stem. Auxin leads to remeristemisation of permanent cortical tissues and organ differentiation leading to root formation.
(b) On Petiolar abscission:
Experiment:
Leaves of some suitable plants (Ervatamia, Coleus, Phaseolus, etc.) are debladed, i.e., lamina is removed from the petiole and at the cut end of the petioles- auxin paste (0-25%) is applied keeping a few debladed untreated plants as control. Observation is made after a week.
Observation:
It is observed that petioles of the untreated plants abscised while the treated petioles do not (slight but uniform pressure is to be applied to ensure abscission of petioles).
ADVERTISEMENTS:
Inference:
It is the property of auxin to inhibit abscission. Some of the important reasons are petiolar growth, higher auxin gradient at the petiolar end and prevention of mobilisation of metabolites.
(c) On apical dominance:
Experiment:
ADVERTISEMENTS:
It is a common finding that when terminal bud of a shoot is excised, one or more lateral buds promptly develop below. The nature of the influence exerted by the terminal bud on suppressing the growth of the lateral buds is called apical dominance.
Potted sunflower or Coleus plants are taken for this experiment and from some plants the apical buds are removed keeping other plants as control. To half of the excised plants auxin in lanolin paste (0.25%) is applied. Observation is made after 15 to 20 days.
Observation:
Lateral buds come out in plants only where the terminal buds have been removed. Whereas few or no lateral buds develop in auxin-treated and in control plants.
ADVERTISEMENTS:
Inference:
The terminal bud inhibits or suppresses the growth of lateral buds thus establishing dominance over the remaining buds which remain inactive. As soon as the terminal bud is excised the dominance of the terminal bud is removed causing the hitherto suppressed lateral buds to come out.
The effect is thought to be due to the inhibitory property of auxin produced at the terminal bud which is transported downward through the stem. This is further proved by the substitution of the terminal bud with auxin (in lanolin paste) which suppresses again the lateral bud- growth in much the same way as the terminal bud.
Experiment # 2
Demonstration of Effect of Kinetin on Growth:
(a) On cell division:
ADVERTISEMENTS:
Experiment:
1 to 5 ppm kinetin solution is prepared in distilled water. Tobacco pith tissue or carrot tissue is grown in 6 petridishes containing White’s nutrient medium prepared as follows:
White’s nutrient medium:
Each of the following solutions is separately prepared in a minimum volume of water and then the solutions are mixed one by one in the serial order as arranged below with’ constant shaking, otherwise precipitation may appear. The final volume is made up to 1 litre. Before use, ten times dilution of the stock solution is necessary.
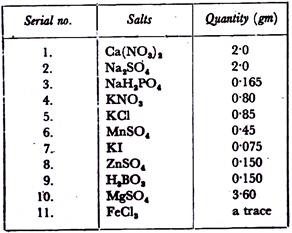
5 ml of 1 ppm, 2 ppm, 3 ppm, 4 ppm, and 5 ppm kinetin solutions are added to the first, second, third, fourth and fifth petridishes respectively. One petridish is kept as control where instead of kinetin solution, 5 ml distilled water is added. Observations are made after seven days.
ADVERTISEMENTS:
Observation:
Cell formation is noted in all the treatments (the area of the cell tissue or the per cent of new cells formed may be measured). Maximum cell formation has been observed in the concentration range of 1 to 2 ppm kinetin.
Inference:
Kinetin induces cell division. The adenine moiety of kinetin molecule appears essential for this process. The side chain of kinetin may also possess some physical property for regulating the efficiency of it to induce cell division.
N.B. Not only does kinetin promote cell division, but also induce cell enlargement. Kinetin treatment of leaf discs cut from etiolated leaves, e.g., of Phaseolus sps causes significant cell enlargement. The effect of kinetin on cell enlargement is usually associated with IAA and gibberellins.
ADVERTISEMENTS:
(b) On leaf senescence:
Experiment:
Vigna or Phaseolus seedlings are most suitable for this experiment. 25 ppm kinetin solution is applied on both surfaces of one of the primary leaves leaving the other as such. The trifoliate leaves are removed as soon as they appear. Observation is made from time to time.
Observation:
It is observed that kinetin treated leaf remains green and fresh while the untreated opposite leaf of the pair becomes yellow.
Inference:
Kinetin is a potent arrestor of senescence. Leaves treated with kinetin draw metabolites from untreated region and also cause an increase of protein and nucleic acid synthesis. Since kinetin is relatively immobile it is not easily transported to the untreated regions. Hence kinetin treated leaf remains green due to deferal of senescence while the untreated leaf shows the sign of senescence much earlier.
Experiment # 3
Demonstration of effect of Gibberellin on Growth:
(a) On seedling growth:
Experiment:
Several suitable seedlings of Vigna or Phaseolus are grown in two petridishes. One lot of seedlings is sprayed with 20 ppm gibberellic acid (GA3) solution with the help of an atomiser or a sprayer. The other lot is sprayed with distilled water which serves as a control. Observations are made after one week by measuring the height of the seedlings.
Observation:
It is observed that the height of the gibberellin-treated seedlings has increased considerably compared to that of control seedlings.
Inference:
Gibberellin, like IAA, promotes cell elongation in the intermodal region and thus increases seedling growth. The rate of increase is much higher in case of gibberellin than in IAA.
(b) On Light-inhibited stem growth:
Experiment:
Three lots of Vigna or Phaseolus seedlings are grown in three petridishes. One lot is kept in dark and the other two lots are kept in light of which one lot is sprayed with 20 ppm gibberellic acid (GAg) solution. The stem growth of the seedlings in each case is measured and compared.
Observation:
It is observed that height increases considerably in case of dark-grown and light-grown but gibberellin-treated seedlings compared to only light-grown seedlings.
Inference:
Light has an inhibitory effect on stem elongation. Application of gibberellin to the plants growing in light greatly increases their stem growth. Reversal of light induced inhibition of stem elongation by application of gibberellin suggests that endogenous gibberellin is the limiting factor in stem growth.
It may be concluded that light causes inhibition of stem growth by lowering the level of endogenous gibberellin and this inhibition is erased by applying exogenous gibberellin to the plants.
Experiment # 4
Inhibition of Protoplasmic Movement:
Experiment:
The protoplasm of most living plant cell is frequently in active motion. Sections of Chara, Vallisneria, Elodea, etc. show this movement. Here the granular plastids can be seen to move along this protoplasmic stream (cyclosis).
Some sections of Vallisneria or Elodea or Chara are put to solutions of respiratory inhibitor like malonic acid, cyanide or un-coupler of ATP like DNP in dilute concentrations. These treated sections are compared with untreated sections.
Observation:
It is observed that the rate of movement of plastids or the protoplasmic streaming decreases considerably in case of inhibitor treatment in comparison to controls.
Inference:
Though the mechanism of protoplasmic movement is not known yet, the involvement of respiratory energy for this movement is clearly indicated-from studies with respiratory inhibitors.
Experiment # 5
Inhibition of Water Uptake:
Experiment:
Slices of peeled potato tuber of uniform thickness are prepared. Equal weights of slices are taken in two petridishes, one containing distilled water and the other containing any one of the inhibitor solutions mentioned in the above experiment.
The weights of the slices of both the petridishes are taken at an interval of fifteen minutes for a period of two hours. The slices should be blotted carefully before taking weights.
Result:
The weighty of slices kept in both the petridishes increased gradually but the rate of increase of water uptake is much higher in distilled water medium than that of slices kept in inhibitor solutions.
Discussion:
Some amount of water is absorbed by the plant cells by active process, i.e., at the expense of respiratory energy. Treatment of the plant tissue with metabolic inhibitor will affect the process thereby reducing the rate of absorption.
N.B. The active absorption of ions can also be inhibited by using metabolic inhibitors or inhibitors of protein synthesis (chloramphenicol) indicating that this process is dependent upon energy expenditure and enzyme activity.
The active accumulation of potassium ions by Nitella can be studied by keeping the algae in a dilute solution of KNO3 as well as by keeping them in the KNO3 solution containing a metabolic inhibitor. The more potassium ions will be accumulated by the algae, the less will be the conductivity of the external medium which can be detected by a Conductivity meter.
Experiment # 6
Inhibition of Transpiration:
Experiment:
Two transpiration experiments are set up as in Expt. 8(c) of Chapter VI. In one conical flask, instead of water, some inhibitor solution of either respiration (malonic acid) or protein synthesis (chloramphenicol) or un-coupler of oxidative phosphorylation (DNP) or any other antitranspirant like abscisic acid is taken. The rate of transpiration in both the cases is determined.
Results:
The rate of transpiration in control set is found to be higher than that of the inhibitor-treated set.
Discussion:
Transpiration is dependent upon opening and closing of stomata and the phenomenon of opening and closing of stomata is again an energy-driven enzyme catalyzed process. Thus the rate of transpiration is always inhibited by a respiratory inhibitor which has an effect on oxidative phosphorylation and by inhibitor of protein synthesis which has an effect on enzyme synthesis and activity.
Experiment # 7
Inhibition of Respiration:
Experiment:
Two experiments on respiration are set up as in Expt. 1 of Chapter X. Two lots of germinating gram seeds of equal weights are taken.
One lot of such seed is treated with a dilute solution of any respiratory inhibitor (iodoacetic acid or iodoacetamide or fluoride which are inhibitors of glycolysis; or cyanide, malonic acid, etc. which are inhibitors of Krebs cycle) for 2 hours and the other lot is similarly treated with distilled water only. The rates of respiration in both the lots are measured.
Results:
The rate of respiration is much lower in inhibitor-treated seeds than the untreated control.
Discussion:
Respiratory inhibitors inhibit enzyme-catalysed reaction as a consequence of which the rate of respiration is inhibited.
Iodoacetamide or iodoacetic acid prevents oxidation of triosephosphate to phosphoglyceric acid, fluoride inhibits the enzyme enolase, cyanide inhibits the activity of oxidase enzyme and malonic acid inhibits the enzyme succinic dehydrogenase.
Experiment # 8
Inhibition of Photosynthesis:
Experiment:
In one beaker, instead of water, weak cyanide solution (inhibitor of dark reaction) or DCMU or CMU solution or atrazine solution (inhibitors of photochemical reaction) is taken.
The rate of photosynthesis is measured in both the cases under moderate and uniform light intensity. Other set is kept as control.
Results:
The rate of photosynthesis is found to be less in inhibitor treated plants compared to control.
Discussion:
Cyanide is an inhibitor of dark reaction, i.e., reductive fixation of CO2 which is cyanide sensitive. DCMU or CMU inhibits O2 evolution by chloroplast, i.e., photolysis of water in photosystem II and atrazine inhibits phosphorylation in photosystem I and hence the reduction in the rate of photosynthesis.
Experiment # 9
Inhibition of Growth:
Experiment:
Some seedlings of Phaseolus or Vigna arc grown in two earthenware pots. 50 ppm solution of any growth retardant (maleic hydrazide, AIar-85, CCC, AMO-1618, etc.) is applied to 10 seedling of one pot every alternate day for a period of 10 days.
The seedlings of the other pot are kept as control. The growths in height of the seedlings as well as leaf areas of the seedlings are measured in treated and untreated controls.
Results:
The average height and the leaf area are recorded in treated and untreated controls and results are compared.
Discussion:
The growth in height of the seedlings and area of the leaves are much retarded in case of inhibitor-treated seedlings as compared to untreated controls. These growth retardants mainly act antagonistically with the growth hormones particularly with the gibberellins thereby reducing the growth of the plant as a whole.
Experiment # 10
Inhibition of Germination:
Experiment:
Some suitable non-dormant and viable seeds are taken. Two filter papers are taken in two petridishes. To one petridish 5 ml of any germination inhibitor (coumaric acid or coumarin, abscisic acid, cycloheximide, ferulic acid, etc.) of 10 to 20 ppm concentration is taken.
To the other 5 ml of distilled water is added. Twenty seeds are arranged on the filter paper of each petridish and the number of seeds germinated every day is noted for a period of one week.
Results:
The speed of germination (number of seeds germinated each day) and the final percentage of germination are recorded in each case.
Discussion:
Germination is inhibited by treatment with inhibitors. The inhibitors mainly act on the enzyme system, particularly hydrolases, which takes active role in germination process by breaking down complex substances into simpler ones. These inhibitors may also act antagonistically on gibberellin action which is essential for germination.
Experiment # 11
Inhibition of Flowering:
Experiment:
Seedlings of Lactuca or Nitiana are grown in pots. Some seedlings are treated with 50 ppm of any anti-gibberellin (AMO-1618, Phosphon-D, CCC, etc.) every alternate day for a period of 10 days. Some seedlings are kept as control.
Results:
The days required for bolting (emergence of the inflorescence stalk) is recorded in treated and untreated controls.
Discussion:
Gibberellin has an active role in flowering of long day rosette plant. So flowering is inhibited in case of long day rosette plants by application of anti-gibberellins. However, the mechanism of action of these anti-gibberellins is not yet clear.
Experiment # 12
Bioassay of Natural Growth Regulators:
Assay is a term originally employed for the determination of the proportion of a metal in an ore or an alloy. In recent years it has been applied by biologists to the determination of specific activity of such biological products as hormones, inhibitors, vitamins, antibiotics, etc.
The main assay methods which have been employed in determination of growth substances are given below. The details of the methods will be available in any standard text book on growth substances.
a. Bioassay of IAA:
(i) Coleoptile curvature test by Avena coleoptile.
(ii) Other curvature tests with cylindrical organs in which cotyledons are removed by an oblique cut from the tops or hypocotyls of Cepfudaria sp. and Raphatms sativa.
(iii) Curvature test with split organs or split pea curvature test with young stem of the garden pea grown in dark for about a week (etiolated).
(iv) Segment straight growth test with Avena coleoptile.
(v) Root growth test with Triticum vulgare, Lens esculantus, etc. Epinastic test with tomato leaf.
b. Bioassay of Cytokinins:
(i) Callus tissue growth with Nicotiana tabacum stem pith, soybean cotyledons, and carrot tissue.
(ii) Bud formation with protonema of moss.
(iii) Leaf expansion with Phaseolus vulgaris, radish, etc.
ADVERTISEMENTS:
(iv) Leaf yellowing test (delay of senescence) with Xanthium, barley, oat and bean leaves, radish and cabbage leaf discs.
c. Bioassay of Gibberellins:
(i) Seedling test with dwarf pea, dwarf maize, seedlings of lettuce, etc.
(ii) Leaf growth with segments of immature first leaf base of certain cereals, e.g., Avena, Triticum, Hordeum, etc.
(iii) Barley endosperm test with barley endosperm and a-amylase activity.
(iv) Leaf senescence test with isolated leaf discs of Taraxacum sp., Rumex sp. on chlorophyll retention.
d. Bioassay of Abscisic Acid:
(i) Acceleration of abscission in excised abscission zone.
(ii) Inhibition of coleoptile curvature or straight growth.
(iii) Inhibition of seed germination, including growth of excised embryos.
(iv) Inhibition of growth of rice seedling.
e. Bioassay of Ethylene:
(i) Expinasty of leaves
(ii) Swelling of Stems and
These three responses are together known as “triple response”. Etiolated pea seedlings are suitable materials for ethylene bioassay.