
ADVERTISEMENTS:
The following points highlight the top five characteristics of pollen used in taxonomy. The characteristics are: 1. Shape, Size and Symmetry of Pollen 2. Pollen Association 3. Pollen Nuclear Number 4. Apertural Forms 5. Exine Stratification.
Characteristic # 1. Shape, Size and Symmetry of Pollen:
Size and shape of pollen are important diagnostic characters. The shape may be circular, triangular, etc., in polar view, whereas it may be per-oblate, spheroidal, sub-spheroidal, sub-oblate, oblate, oblate spheroidal, pro-late, pro-late spheroidal, sub-prolate, per-prolate, etc. in equatorial view (Fig. 8.19).
There is a considerable variation in the size of pollen in different plants. In modern flowering plants, the size of the pollen grains varies from about 5×2.4 µm to 200 nm or slightly more in certain members of Cucurbitaceae.
ADVERTISEMENTS:
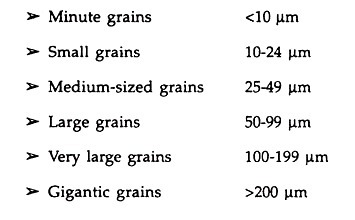
Several size Classes, have been recognized depending on the length of pollen, which are as follows:
Regarding symmetry, they may be symmetric or asymmetric (Fig. 8.18). Symmetric pollen as a rule may be either radially or bilaterally symmetrical. The bilateral pollen further may be planoconvex, concavo-convex or biconvex in lateral view.
Characteristic # 2. Pollen Association:
Pollen associations, such as their arrangement in tetrads, presence of pollinia, etc. are characteristic of several angiospermic families. They are seen in diverse families of the dicotyledons, such as Annonaceae, Droseraceae, Mimosaceae, Monimiaceae and Winteraceae.
However, in the monocotyledons, they are restricted to such advanced families as the Cyperaceae and Juncaceae. Extreme cohesion of pollen in the form of pollinia is seen in the Asclepiadaceae and Orchidaceae.
These associations can be of the following types (Fig. 8.20):
a. Monads — Single pollen grains which when mature, do not remain united with other pollen grains e.g. Poaceae.
b. Dyads — Pollen grains when united in pairs e.g. Scheuchzeria palustris.
c. Tetrads — When four pollen grains are united together.
They may be further categorized into the following types:
d. Tetrahedral — One the pollen grains lie centrally over the other three e.g., Drimys.
ADVERTISEMENTS:
e. Tetragonal (isobilateral) — All pollen grains lie in one plane in a bluntly quadrangular manner e.g., in monocotyledons.
f. Rhomboidal — e.g., Anona muricata.
g. Decussate (cross) — The pollen grains lie pair-wise at right angles to each other e.g., Magnolia grandiflora.
h. T-shaped — In this case, the first division of the PMC is transverse and a dyad cell is formed of which either the upper or the lower dyad cell instead of undergoing a transverse division, divides vertically or longitudinally, yielding either a straight, or inverted T-shaped configuration e.g., Aristotolochia polyanthus.
ADVERTISEMENTS:
i. Linear — In this case, the first division is always transverse, giving rise to two dyad cells. The second division is also transverse, which results in a linear tetrad e.g., Mimosa pudica.
a. Pseudomonad (crypto tetrad) — It is a degraded tetrad in which monad-like tetrads are formed without dividing walls between the four components. One nucleus is normally developed, whereas the remaining three are obliterated e.g., Cyperaceae.
b. Massula (ae) — They are also called polyads. In this case each of the tetrad cells divides once or twice resulting in a massulae of 8-64 cells, which remain together after maturity and are held together in small units e.g., Mimosaceae.
c. Pollinium (ia) — This includes a unit of all grains produced by one theca (anther loculus). The pollinium apparatus is the functional unit of a ‘corpusculum’ with its two attached arms and pollinia e.g., Cryptostegia, Calotropis.
ADVERTISEMENTS:
The taxonomic usefulness of pollen association in the multipalynous family Orchidaceae has been pointed out by Vij and Kashyap (1978) who studied 50 species under Orchidaceae and observed that single pollen grains were found in the relatively primitive groups (Cephalanthera ensifolia, Cypripedium cordigerum, Paphiopedilum villosum), tetrads in the tribe Neottieae and Epidendrieae and perfect massulae in the Orchideae and some Neottieae, where the shape of the massulae can be of taxonomic value for lower-level classifications.
Characteristic # 3. Pollen Nuclear Number:
ADVERTISEMENTS:
The number of nuclei in the pollen grains at the time of their release is a valuable taxonomic and phylogenetic character. The number of nuclei in a pollen grain may be regarded as a function of the stage at which the pollen grains are released. In the angiosperms, the pollen show a definite pattern at which stage a pollen is released, and they may be released either:
i. At the two-nucleate stage, before the division of the generative nucleus, or
ii. At the three-nucleate stage, after the division of the generative nucleus.
Further it has been found that:
iii. The three-nucleate pollen grains have arisen from the two-nucleate ones.
iv. The change is irreversible.
ADVERTISEMENTS:
The stage at which the pollen are released varies in different groups of angiosperms:
Dicotyledons :
The pollen nuclear number do not show a definite pattern.
a. Certain orders have consistently tri-nucleate pollen grains, e.g. Asterales, Caryophyllales, Plumbaginales, Polygonales.
b. Some orders are heterogenous in having both two-and three-nucleate pollen and both the types may even occur in one and the same genus, as has been observed in Burmannia, Ipomoea and Lobelia.
Monocotyledons :
ADVERTISEMENTS:
In the Monocotyledons, the highly reduced wind-pollinated groups, aquatics and achlorophyllous saprophytes show the three-nucleate pollen grains.
Characteristic # 4. Apertural Forms:
The form, number, distribution and position of apertures are important palynological criteria in assessing relationships and phylogeny of plants. Almost all palynological discussions on plant relationships and phylogeny are based on the form, number, distribution and position of the apertures.
Apertures are generally thin-walled areas in the outer pollen wall or exine, through which the pollen tube usually emerges at the time of germination.
There are two basic forms of apertures:
Simple Apertures :
Apertures are either long or short and rounded and may be of following types (Fig. 8.21):
Sulcus :
It is an elongated latitudinal ectoaperture situated at the distal or proximal pole of a pollen grain. A sulcus has the same shape as a colpus, but differs in orientation [e.g. Nymphaea (Nymphaeaceae), Sparganium (Typhaceae)]. Sulcate grains are either:
a. Anasulcate (distal)
b. Catasulcate (proximal)
c. Zonosulcate (extend right around the grain)
Ulcus :
A more or less rounded aperture situated at the distal or proximal pole of a pollen grain is termed ulcus and may be derived from a sulcus or a sulcoid aperture [e.g. Sparganium (Sparganiaceae), Typha (Typhaceae)].
Colpus :
It is an aperture, which is long-furrow-like i.e. twice as long as broad (with a length and breadth ratio greater than 2) and the grain is referred to as colpate. Two basic types of pollen occur in the angiosperms:
a. Monocolpate — It is a characteristic of the monocotyledons and some dicotyledonous families such as woody Ranales, and have a single furrow on one side of the pollen remote from the point of contact in the tetrad.
b. Tricolpate — It is a characteristic of most of the dicotyledons, and are provided with three meridionally placed furrows.
Two other types of pollen also occur, which are considered to have been derived from both the monocolpate and tricolpate types as a result of specialization.
They are:
a. Acolpate -They are pollen without a furrow.
b. Pantocolpate – They are pollen with many furrows.
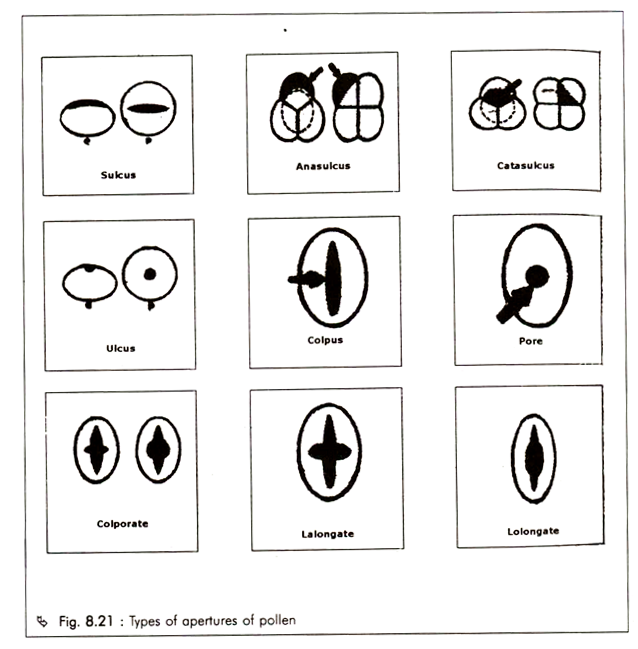
Pore:
It is a circular or elliptic aperture is short with length and breadth ratio less than 2 and the pollen grain is called porate [e.g. Myrica gale (Myricaceae)].
Compound Apertures:
They are apertures consisting of an outer or marginal colpal (furrow-like) part and an inner or central distinctly delimited oral part. Ora are either circular or lalongate i.e., transversely elongated endoaperture [e.g. Filipendula (Rosaceae)] or lolongate i.e., longitudinally elongated endoaperture [e.g. Rumex spp. (Polygonaceae)]. In relation to ora, the furrows or apertures will be named colporate or perorate.
The position of the apertures varies in different groups of plants (Fig. 8.22) and may be:
Proximal:
Found in uniaperturate forms and in terms of evolution are the most primitive. They are found in the gymnosperms and monocotyledons.
Zonal:
Found in multiaperturate forms and are found in the dicotyledons. In this case the furrows or pores are situated along the equatorial line.
Global:
Found in multiaperturate forms and are found in the dicotyledons. In terms of evolution, they are the most advanced. The furrows or pores are distributed over the entire surface in this type.
Most angiospermous pollen grains fall into the following (Fig. 8.23) two general types:
Monoaperturate:
They are characteristic of the monocotyledons and some members of Magnoliidae. Uniaperturate pollen grains have a single germinal furrow or pore situated at the proximal or distal position.
Triaperturate:
The triaperturate or triaperturate-derived pollen grains occur in most of the dicotyledons and are unknown elsewhere. These pollen grains have three germinal furrows, either radiating like the lines of a trilete mark from a common point or forming a triangle at the distal position. Some less common types have been derived from the triaperturate form.
Various other forms of pollen which appears to have evolved from the uniaperturate and triaperturate forms (Fig. 8.23), include:
Multiaperturate:
The multiaperturate type results from cross-partitioning of one germinal furrow into several. It is found in both monocotyledons and dicotyledons.
Diaperturate:
The diaperturate type also results from cross-partitioning and is found in certain monocotyledons.
Nonaperturate:
The non-aperturate type results from progressive shortening and eventual elimination of the furrow and is found in both monocotyledons and dicotyledons.
The various types of apertural morphoforms provide one of the best taxonomic criteria in routine taxonomic work, being often constant, not very variable and easily visible under a microscope.
Nair has even proposed a palynological classification of the whole plant kingdom based on such data and have classified the plants into:
(i) Amorphosporophyta – includes thallophytes with no apertural markings.
(ii) Trimorphosporophyta – includes Archegoniatae with monolete, trilete or alete pollen.
(iii) Polymorphosporophyta – containing angiosperms with diverse forms of apertural types.
There is a remarkable relationship between morphological types of pollen and taxonomic subdivisions in angiosperm families. An example is illustrated by the family Lecythidaceae.
Based on palynological data Miiller (1972) found that the pollen grains of its members fall into two groups, tricolpate and syntricolpate, and that in combination with other characters this can effectively be utilized in subdividing the family. Apertural characters have also been used in lower level taxonomy.
For example in the case of Sesamum, the number of colpi is species-specific i.e., eleven in S. indicum, eight in S. laciniatum and nine in S. prostratum.



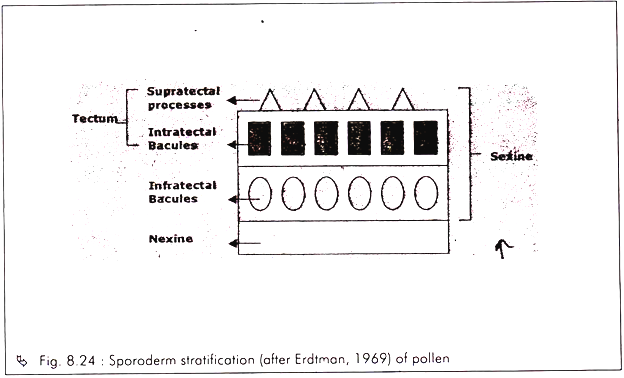
Characteristic # 5. Exine Stratification:
The pollen wall or sporoderm is generally layered (stratified). In angiosperms, it consists of two layers – the outer resistant layer composed of sporopollenin, known as exine and the inner more or less cellulose layer called intine.
Topographically, the exine consists of two well-defined sub-layers – the outer sculptured sexine or ektexine and the inner non- sculptured nexine or endexine (Fig. 8.24).
The sexine further consists of two main parts – a partially or wholly covering layer called tectum below which, are the rods or rod like elements called bacules. According to Walker, there are three basic types of pollen based on the characteristic of tectum.
They are:
a. Tectate — have a continuous tectum. Tectate pollen may be either
i. Tectate-perforate
ii. Tectate-imperforate
b. Semitectate — tectum is not continuous.
c. Intectate — without an apparent tectum.
The trend of exine evolution is from tectate-imperforate to tectate-perforate to semitectate and, more rarely, to intectate pollen. The sculpturing of the external substance (exine) of pollen grains presents a remarkable series of microscopic characters and may vary from family to family.
Commonly, the family and often the genus may be identified by the pollen grains. In general, angiosperm pollen grains have a massive exine and a thin intine.
However, certain taxa among Monocotyledons are unique in having a highly reduced exine and an elaborate intine e.g. Amaryllidaceae, Cannaceae, Costaceae, Heliconiaceae, Iridaceae, Marantaceae, Musaceae, Strelitziaceae and Zingiberaceae. Further the pollen in anemophilous group is smooth and thin, whereas the ones distributed by insects or -birds, are characterized by various types of sculpturing on the exine.

The ektexine or sexine may be either of the following three types (Fig. 8.25):
Columellar :
A rod-like element of the sexine/ectexine, either supporting a tectum or a caput, e.g. Angiosperms
Alveolar :
Describing a type of sexine/ectexine structure, in which the infratectal layer is characterized by partitions forming compartments of irregular size and shape, e.g. Gymnosperms (Pinaceae)
Granular :
A type of exine stratification in which the infratectal layer is composed of more or less rounded, granules rather than of columellae or other structures, e.g. some Gymnosperms and Angiosperms
The endexine or nexine may be either of the following two types:
a. Laminated — e.g. Gymnosperms
b. Non-laminated — e.g. Angiosperms
In recent years the pollen wall structure has become the subject of very detailed and critical study, employing phase contrast and ultraviolet microscopy as well as the more conventional light microscopy.
Ultrafine sections of grains have also been studied under the electron microscope. Ultraviolet microscopy has proved useful in the study of fine details of the intine and cytoplasm. With the improvement of techniques it has been possible to demonstrate fine details of the structure of the walls (sporoderms) and their stratification.
The exine ornamentation may be of either of the two types (Fig. 8.26):
Depression type :
In this case the exine is smooth. This includes many subtypes, such as
i. Areolate — luminoid network surrounds islands of raised areas i.e., a feature of ornamentation in which the exine is composed of circular or polygonal areas separated by grooves.

ii. Foveolate — A feature of ornamentation consisting of more or less rounded depressions or Lumina more than l^m in diameter. The distance between foveolae is greater than their breadth.
iii. Fossulate — A feature of ornamentation consisting of an elongated, irregular groove in the surface.
ADVERTISEMENTS:
iv. Rugulate — A type of ornamentation consisting of elongated sexine elements more than 1 µm long, arranged in an irregular pattern that is intermediate between striate and reticulate.
v. Reticulate — Lumina in the form of a network-like pattern consisting of Lumina or other spaces wider than 1 µm bordered by elements narrower than the Lumina.
vi. Retipliate — incomplete fusion of columella i.e., describing a reticulum formed by rows of pila instead of muri.
vii. Scrobiculate — A rounded or elongate tectal perforation, less than 1 µm in length or diameter.
viii. Striate — elongated, generally parallel elements separated by grooves.
Excrescences type:
In this case the exine carry processes or excrescences of various kinds. This includes many subtypes, such as
I. Baculate — cylindrical or rod shaped free-standing excrescences, more than 1 µm in length and less than this in diameter.
II. Clavate — club shaped excrescences that is higher than 1 µm, with diameter smaller than height and thicker at the apex than the base.
III. Gemmate — excrescences are in the form of rounded warts with constricted base, higher than 1 µm, and that has approximately the same width as its height.
IV. Granulose — A type of exine stratification in which the infratectal layer is composed of more or less rounded, granules rather than of columellae or other structures.
V. Spinose — long pointed excrescences
VI. Spinulose — excrescences are as long and tapering pointed elements, exceeding 1 µm.
VII. Tuberculate — excrescences in the form of knobbly projections.
VIII. Verrucate — excrescences are rounded but base is not constricted i.e., wart-like excrescences, more than 1 µm wide, that is broader than it is high and is not constricted at the base.