
ADVERTISEMENTS:
The following points highlight the top eight types of Gene-Gene Interactions. The types are: 1. Interaction between Dominant Factors 2. Complementary Factors (9:7 Ratio) 3. Supplementary Factor (9; 3: 4 Ratio) 4. Epistasis(12:3:l Ratio) 5. Effect-of Inhibitory Factor (13: 3 Ratio) 6. Duplicate Factors (15: 1 Ratio) 7. Multiple Factors or Polygenic Inheritance 8. Lethal Factor (2:1 Ratio).
Gene-Gene Interactions: Type # 1. Interaction between Dominant Factors:
Some of the heritable characters in plants and animals depend upon the combined effect of two or more pairs of dominant genes.
Bateson and Punnett have explained it by taking example of inheritance of comb types in fowls.
ADVERTISEMENTS:
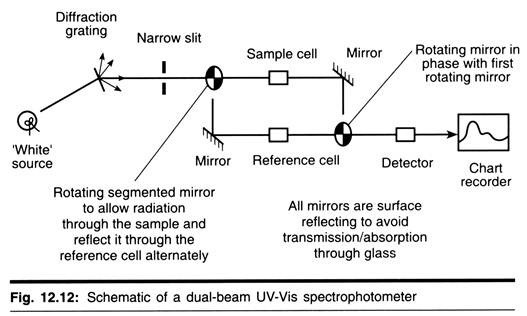
The following four main types of combs (Fig. 15.3) are common among fowls:
(i) Rose comb,
(ii) Pea comb,
(iii) Walnut comb, and
ADVERTISEMENTS:
(iv) Single comb.

Among these, pea and rose types are governed by two separate dominant genes which are found on separate chromosomes and single comb is recessive to all of them.
If the dominant gene for a comb is represented by ‘P’ and that of Rose comb by ‘R’ the genotypes of the above three homozygous forms will be as follows:
Pea = PP rr
Rose = ppRR
Single = PP rr
When Rose and Pea combed fowls were mated, progeny with a new type of comb, called the Walnut comb (genotype Pp Rr) appeared in the F2. In the production of walnut comb, therefore, two independent dominant factors ‘P’ and ‘R’ are jointly responsible.
When these F1, hybrids were inbred, they produced all the four types of combs in the typical dihybrid ratio 9 Walnut: 3Rose:3Pea:1 Single. This is shown in Fig. 15.4.

Gene-Gene Interactions: Type # 2. Complementary Factors (9:7 Ratio):
Certain characters are produced by the interaction between two or more genes inherited from different parents. These genes are complementary to one another, i.e., if present alone these genes remain unexpressed, but are expressed only when they are combined in F1, through suitable crossing.
W. Bateson and R.C. Punnett crossed two white flowered varieties of sweet pea (lathyrus odoratus). In the experiment all the plants of F1, generation developed red flowers instead of expected white flowers. These F1, plants, on self-pollination, produced red and white flowered plants in the ratio 9: 7 in F2 generation.
Bateson explained his Sweetpea experiment in the following way:

Summary:
ADVERTISEMENTS:
Red flowered plants with both complementary genes C and R = 9, White flowered plants having either ‘C’ or ‘R’ = 7.
From the checker board it is clear that the red colouration of the flower is due to interaction of two complementary factors C and R. The parents developed white flowers because they lacked either gene ‘C’ or gene ‘R’ in their genotypic expressions CC cc and RR rr. F1, hybrids received both the complementary genes C and R, hence all the flowers in that generation were red.
The ratio 9: 7 obtained in F2 generation is actually a modification of normal dihybrid ratio (9:3:3: 1).
ADVERTISEMENTS:
Similar cases are reported in many other plants. In Sorghum (jowar), for example, there are two white grained races which, when crossed, produced F1, hybrids with brown grains. When brown grained F1 plants were self-pollinated, they produced brown grained and white grained plants in the ratio 9:7.
In paddy, the plants of two varieties with colourless stigmas, when crossed, produced F1, hybrids with purple stigma. By selfing the F1, hybrids, plants with purple and white stigmas were obtained in the 9: 7 ratio in F2.
Gene-Gene Interactions: Type # 3. Supplementary Factor (9; 3: 4 Ratio):
Sometimes interaction involves two different types of genes in which one type can express itself independently but other has no expression of its own. When two such genes interact, they produce a new character In Sorghum (Jowar) the black purple colour (P) is dominant over brown (p). Blackish pigmentation is found to change to red in certain crosses.
The modifying factor Q borne on another chromosome has no expression of its own in presence of other recessive factor ‘p’. Similarly, the recessive factor ‘q’ is also without any phenotypic effect but when both dominants P and Q are brought together, the blackish purple colour is changed to red.
ADVERTISEMENTS:
The theoretical results to be expected from a cross between blackish purple and brown glumed variety can be explained in the following way:
An example of similar type is met within rats and rodents. In rats, there are three common skin or coat colours. These are Black, Albino, and Wild type Agouti (yellow hair with black bases). The black coat colour is governed by dominant gene ‘C’ and albino colour by dominant gene ‘A’.
The agouti colour is governed by interaction of two dominant genes A and C. ‘A’ does not show any visible effect if it is alone, but the gene C is able to express itself independently.
When a true breeding black rat is crossed with an albino carrying the dominant gene A, all the F1, hybrids are of the agouti type. When F1 individuals are inbred, the F1, generation is represented by agouti, black and albino in 9: 3: 4 ratios.
These results can be explained in the following way:


Gene-Gene Interactions: Type # 4. Epistasis(12:3:l Ratio):
According to Mendel’s laws of inheritance, when two contrasting characters (allelomorphs) are brought together, one of them dominates over the other in F1. In the phenomenon of epistasis, two independent non-allelic genes affecting the same trait of an individual interact in such a way that one over-masks the expression of the other.
Here the gene which prevents the expression of another non-allelic gene is said to be epistatic and the suppressed gene is said to be hypostatic.
In cholam (sorghum caudatum), the pearly grain colour is dominant over chalky. Pearly colour develops when dominant gene ‘Z’ is present, and chalky colour develops in absence of it. Another dominant gene ‘W which is responsible for the red colour of seeds, is dominant over the gene Z.
When ‘ W’ is present, the grain will always be red irrespective of the presence of other gene Z. In the absence of W, the grain colour will be pearly or chalky depending upon the presence or absence of gene Z.
Thus the genotypes of all the coloured grained plants will be as under:
ADVERTISEMENTS:
Red grained = WWzz or WWZZ
Pearly grained = wwZZ
Chalky grained = wwzz
When a red grained type with the genotype WWzz is crossed with pearly variety (wwZZ), F1 offsprings are red grained (Ww Zz). In F, generation three kinds of plants; red, pearly and chalky, appear in the ratio 12:3: 1 which is a modified dihybrid ratio.
The mode of inheritance can be represented in the following way:

In the squashes (cucurbita pepo), Sinnot has shown that there are three common fruit colours, white, yellow and green. Sinnot has shown that white was dominant over both yellow and green and yellow was dominant over green only.
In a cross between white variety (genotype WW YY) and green variety (genotype wwyy), the F, progeny appeared white fruited (genotype Ww Yy) which on selfing gave a 12 white: 3 Yellow: 1 green ratio. Here ‘ W’ acts as an epistatic factor and ‘ Y’ is active only with ‘w’;

From the above experiment, it is clear that each fruit colour is being governed by two non-allelic genes W and Y. If gene W is in dominant condition, dominant Y gene will not show its effect. Gene Y is active only when it is combined with recessive ‘w’. V.
Gene-Gene Interactions: Type # 5. Effect-of Inhibitory Factor (13: 3 Ratio):
In this modification or gene interaction, one dominant factor inhibits or suppresses the phenotypic expression of another dominant gene found on different chromosome. This may be explained by taking example of pigmentation of leaf in rice plants. Here purple colour of the leaf is due to factor Lp and green due to lp. The purple colour is dominant over the green.
There is another factor T. If dominant factor T is present with the factor Lp, the purple colouration (due to factor Lp) is inhibited and the leaf becomes normal green. The factor T has thus no visible effect of its own but it inhibits the colour expression of gene Lp.
In a cross between purple pigmented (ii Lp Lp) and a green (I I lp lp) rice plant, the F1, plants were all green (I i Lplp) and in F2 there appeared green and purple pigmented plants in 13: 3 ratio.
The results are explained below:

A similar effect is seen in the inheritance of plume colours in fowl. White Wyandotte and white leghorn are two breeds of fowls with white plumes. The white plumage of the white wyandottes is recessive to the coloured plumage. The white leghorns possess a colour factor which is prevented from being expressed by an inhibiting gene.
If the inhibiting factor is symbolised by ‘I’ and the colour factor by ‘C, the genotype of white leghorn can be expressed by CCII and that of white Wyandotte by ccii.
When white Leghorns (CCII) and white Wyandotte (ccii) are crossed, the F1, hybrids are all white (Cc Ii). When the F, hybrids are inbred, the offsprings in F2 appear in the ratio 13 White:3 coloured.
The results can be explained in the following way:

Gene-Gene Interactions: Type # 6. Duplicate Factors (15: 1 Ratio):
In this type of interaction two or more genes either individually or in combination produce the same phenotypic effect. An example of this type of interaction is the inheritance of awn in rice grain. In a cross between ‘awned’ and ‘awnless’ types, F1, rice grains were all awned. Awned character is thus dominant over awnless.
The F1, plants on self-pollination give awned and awnless types in 15: 1 ratio in F2 generation. This result can be explained if it is supposed that awned character is governed by two dominant genes, say A1, and A2. The presence of one or both dominant genes results in awned grains and in absence of both these dominant genes awnless grains develop.


From the above cross h is clear that out of sixteen offsprings in the F2 generation, fifteen have one or both the dominant genes, so they are all awned type and only one is such as is double recessive, hence awnless.
The inheritance of chlorophyll in ragi can be cited as second example. Two factors C1, and C2 produced chlorophyll.
When a cross was made between chlorophyllous and achlorophyllous parents, F1, seedlings with factors C1, C2 were chlorophyllous (green) and F2 individuals were green and non-green in ratio 15:1.
Gene-Gene Interactions: Type # 7. Multiple Factors or Polygenic Inheritance:
The term polygenic inheritance is applied when two or more independent pairs of genes affect the same character in the same way and in additive fashion. In such cases, the net effect on the trait depends upon the combined action of several genes, each of which has a small effect on the same trait.
Such genes are called the ‘cumulative genes’ or ‘polygenes’ and the trait influenced by them is known as ‘polygenic trait’. Polygenic traits do not show clear cut difference between individuals and all gradations occur between the two extremes.
The characters are thus quantitative instead of being qualitative, as for example, the height, weight, pigmentation, blood pressure, finger prints, ridge counts and intelligence in man, the amount of fruits and seeds produced by plants and milk or meat produced by animals.
The inheritance of pericarp colour in wheat is polygenic according to Herman, a Swedish Geneticist. Nilsson-Ehle (1909) crossed pure red seeded wheat with pure breeding white seeded wheat. The colour of seed coat is dependent in this case on the genotype of the plant and not the seeds because the pigment is derived from maternal tissue.
The F1, plants were found to have red seed. When selfing was allowed in F, plants, all the 78 F2 plants were found to have red seeds instead of red and white in 3: 1 ratio as expected. He then self-pollinated F2 plants individually and found that 78 plants could be grouped into the following 4 categories on the basis of the ratio obtained in F3 generation.
(i) Group 1:
F2 plants producing red seeded and white seeded plants in a ratio 3:1.
Such plants were eight in number. From this he reasoned that they were heterozygous for one gene pair.
(ii) Group 2:
F2 plants produced 15 red: 1 white; so he reasoned that they were heterozygous for two gene pairs. Such plants were 15 in number.
(iii) Group 3:
F2, plants produced red and white in 63:1 ratio (three homozygous recessive = (1/4)3 = 1/64) and they were 5 in number.
(iv) Group 4:
F2 plants produced only red seeded plants and so he reasoned that they were homozygous for dominant alleles for redness of at least one gene pair.
From these ratios one thing became clear that he was dealing with a triplicate gene situation in which a dominant allele of any one of three separate genes (A,B or C) could cause redness and the original cross must have been AA BB CC (red) x aa bb cc (white) and F1, Aa Bb Cc.
The following Table shows the F2 genotypes and their theoretical probabilities and observed frequencies:


Although Nilsson-Ehle was unlucky not to have picked up the pure white F2 aa bb cc yet his results statistically fit into the triplicate gene model very well.
Nilsson-Ehle actually observed a lot of variations in the amount of redness in red coloured seeds, some being very dark, some being light and some were of intermediate shades and it was difficult for him to make accurate groupings. However, in a dihybrid selfing (say Aa Bb cc x Aa Bb cc), he was able to classify the progeny plants into 5 groups with the following relative frequencies:

To explain the variation in the degree of redness, he hypothesized that the total number of dominant alleles of both gene pairs was determining the degree of colouration. In other words, each dominant gene could be considered to be contributing one dose of redness and the effect of each dose was equal and additive, the more dominant genes a plant inherited darker the colour of pericarp will be.
This idea fits with the observed ratio well forgetting about the third gene pair, although irrelevant here, we can calculate the probability of getting doses of 0,1,2,3 and 4 dominant alleles from the cross Aa Bb X Aa Bb as follows:
Relative frequencies of different dominant doses:
4 dominants—1
3 dominants — 4
2 dominants — 6
1 dominant — 4
0 dominant — 0
The inheritance of fruit shapes in pumpkin (cucurbita pepo) is another example of cumulative effect. A cross between two different races with spherical fruits gives disk shaped F1, offsprings which on self-pollination give 9 disk: 6 sphere: 1 cylindrical in F2 generation.
Another interesting example of the interaction of multiple factors is the inheritance of the skin colour in man. C.B. Devenport (1913) considered that two pairs of genes account for the difference of pigments between negro and concasian (white people).
He conducted his most significant studies in Bermuda and Jamaica where interracial marriages between coloured and white people were relatively frequent and legitimate births were common.
Let us assume that negroes have two pairs of dominant colour genes P1, P1, P2 P2 and whites have only recessive genes P1, P1, P2 P2. A marriage between a negro and a white would give F1, with genotype P1, P1, P2 P2. The individuals of this generation are called mulattoes.
Their colour will be intermediate between two parents. Marriage between mulattoes produces offsprings (F2) ranging in colour from “Pure Negro” through all the intermediate shades to pure white.
The pattern of inheritance can be shown in the following way:

Other investigators have suggested that the range in the skin colours between negro and white is due to more than two pairs of pigment forming genes. Gates (1953) has suggested three pairs of genes. Some workers have suggested the number of colour genes from 2 to 20 pairs.
In Drosophila, eye colour inheritance is governed by multiple alleles. The wild type eye colour is red in Drosophila but the gene for red eye has mutated to white, eosine, cherry wory, apricot and many other colours.
The principle of mulitiple gene inheritance or quantitative inheritance is now considered as most important principle of genetics. It has been strengthened greatly by the use of statistical methods devised by Fisher (1947). Practical applications are being made in plants and animal breeding for the improvement of field quality and colours.
Gene-Gene Interactions: Type # 8. Lethal Factor (2:1 Ratio):
Lethal genes are Mendelian factors whose substitution for their normal allelomorphs results in the developmental or physiological changes and finally causes the death of an organism. The lethal genes may cause death of organism at any time in the life cycle of organism from gamete to adult. Lethal genes may be dominant or recessive, for example, yellow coat colour in mouse is a heterozygous (Yy) character.
Yellow mouse with homozygous dominants (YY) does not survive. Yellow coated mice never breed true. When inbred, these yellow mice always give a progeny of 2 yellow: 1 black instead of monohybrid 3: 1 ratio.
The result of mating between two heterozygous mice may be represented as follows:

The homozygous offsprings (YY) die, because the gene Y in the homozygous condition has lethal effect on the embryo. Thus monohybrid F2 ratio 1:2: 1 is converted into 2: 1 ratio by the death of homozygous yellow (Fig. 15.5). This was verified when spontaneously aborted fetuses were observed in uteri of female.

In fowl, the creeper gene ‘C is lethal in homozygous dominant condition. Creeper fowls with heterozygous genotype (Cc) have short crooked legs. Normals have two recessive genes. When two creepers were mated, a ratio of 2 creepers to 1 normal appeared as shown below:

In human beings a few lethal genes are known which are associated with fatal diseases. Infantile amaurotic idiocy is caused by a recessive allele in homozygous condition. The bearer of this genotype loses its eye sight and becomes mentally degenerated in the early childhood and finally dies.
Colley’s anemia or sickle cell anemia which is common in certain tribes of Africa is yet another example of fatal disease associated with lethal allele.
There are two dominant genes Hb1A and Hb1a. Individuals with genotype Hb1a Hb1s (heterozygous) do not show symptoms of this disease and are phenotypically similar to those with genotype Hb1a Hb1a (homozygous). Individual with genotype Hb1s Hb1s (homozygous) only suffers from the syndrome (disease) sickle cell anemia which leads to death at the adolescence stage.
In this disease, the erythrocytes or red blood cells become distorted and become sickle shaped. These distorted cells block the blood capillaries and impede circulation. Besides, these cells do not help in the exchange of respiratory gases (Fig. 15.6).
In rice, a lethal homozygous recessive (yy) causes the death of seedlings within ten days of growth. Similar homozygous recessive lethal genes have been discovered in cholam, maize and in other Indian crop plants.
In man, several lethal genes are reported to cause diseases which are fatal at different stages of development.
Thalassemia major (anaemia) is an example of such diseases causing death in childhood. Thalassemia minor has very little effect and causes minor abnormalities in the blood.